Общая характеристика скелета животных. Клиническая характеристика детей с различными отклонениями в развитии
Скелет (костяк) - sceletum (рис. 5) - представляет собой пассивную часть аппарата движения. Он состоит из закономерно соединенных друг с другом плотных образований - костей и хрящей; поэтому он служит опорой для мягких тканей и обусловливает форму тела. В основном же скелет - система рычагов, с помощью которых мышцы сохраняют равновесие, обеспечивают передвижение животного в пространстве, относительное перемещение отдельных частей тела или, напротив, фиксируют тело в определенном положении, например при стоянии. Скелет служит вместилищем красного костного мозга (органа кровообразования) и мощным депо минеральных солей.
В дополнение к их очевидной ценности для исследований, связанных с остеопорозами, методы визуализации, используемые в этом исследовании, могут быть применены к любому исследованию скелета. Поскольку прочность кости трудно оценить в клинике, радиологические методы обычно используются для оценки 2 основных детерминант прочности кости: минерализации и структуры. Несмотря на то, что исследователи согласны с тем, что минеральная плотность костной ткани является одним из самых важных факторов прогноза здоровья и силы костей, недавние исследования показали, что эта мера сама по себе может быть недостаточной для определения силы губчатой кости и что трабекулярная архитектура играет решающую роль в определении костной механической свойства и риск перелома.
Различают осевой скелет и скелет конечностей.
Осевой скелет состоит из черепа, или скелета головы, скелета шеи, туловища и скелета хвоста. Скелет туловища, в свою очередь, разделяется на грудной, поясничный и крестцовый отделы. Основная часть скелета шеи, туловища и хвоста - позвоночный столб.
Из всех частей осевого скелета наиболее сложно устроен череп, где размещаются головной мозг, органы зрения, слуха, равновесия, а также начальные отделы пищеварительной и дыхательной трубок - ротовая полость с ротоглоткой и носовая полость с носоглоткой.
Размер и грубая анатомия проксимальной бедра существенно различались среди лабораторных видов, рассмотренных в этом исследовании. Особенности с наиболее очевидными вариациями включают размер головки бедренной кости, длину и угол шейки бедренной кости, размер и морфологию большого вертела, а также форму и размер межстроберной области. Вариация среди видов в трехмерной ориентации шейки бедренной кости относительно диафиза создавала трудности в постоянном позиционировании костных образцов и точном сканировании интересующих областей.
Наиболее полно развит грудной отдел туловища, где, помимо соответствующего участка позвоночного столба, имеются ребра и грудная кость, которые в совокупности формируют грудную клетку. В ней помещаются легкие, сердце и некоторые другие органы. Наиболее слабо у наземных животных развит скелет хвоста.
А. В осевом скелете ясно выступают общие закономерности строения тела животного, одноосность, метамерия и антимерия (рис. 6-А), обусловленные подвижностью животного.
Подобно проксимальной бедренной кости человека, области бедренной головки и шеи проксимальной бедра от собак и макак циномолгуса содержат значительное количество губчатой кости. Результаты экспериментального исследования показывают, что трабекулярная сеть в проксимальном бедре имеет довольно постоянную толщину для всех видов, что позволяет сделать вывод о том, что трабекулярное разделение и связывание способствует изменению числа трабекулов среди видов. Результаты в собаках и макаках из киномолгуса из этого исследования подтверждают более ранние данные о том, что на проксимальной бедрах, где преобладает губчатая кость, необходимо измерять комбинацию минеральной плотности объемной кости в соответствии с структурными параметрами трабекулы, в частности толщины и количества трабекулы, поскольку эти два параметра, используемые в комбинация лучше предсказывает биомеханические свойства проксимальной бедра, а не только один параметр.
Одноосность - monaxonia, или биполярность, выражается в том, что все отделы осевого скелета расположены на одной оси, причем на переднем, головном, полюсе ее находится череп, а на противоположном полюсе - хвост. Признак одноосности позволяет установить в теле животного два направления: краниальное - в сторону головы, или черепа, и каудальное - в сторону хвоста.
Несмотря на незначительное изменение минеральной плотности губчатой и кортикальной костей на шейке бедренной кости и голове, значения плотности были одинаковыми среди видов. Несколько более высокие значения у мышей и крыс могут быть вызваны меньшим количеством, но более толстыми трабекулами у грызунов по сравнению с собаками и нечеловеческими приматами. Как и ожидалось, более крупные виды имели более толстую кору, чтобы поддерживать размерность органов с точки зрения длины и толщины кости и обеспечить соблюдение механических требований, предъявляемых к скелету.
Антимерия, или двусторонняя симметрия, - antimeria (an-ti - против) - характеризуется тем, что скелет, как и тело животного, может быть разделен только одной медианной, или средней сагиттальной, плоскостью на две (правую и левую) симметричные половины одна против другой- антимеры, в то время как любая другая сагиттальная плоскость будет делить тело на неравные, асимметричные половины. В соответствии с этой закономерностью одни части тела, например конечности (правые левые), являются парными, и в них отсутствует симметрия строения (асимметричные кости); другие же части тела, будучи непарными (например,туловище содержит непарные кости- позвонки, грудная кость), построены симметрично. Антимерия дает возможность различать в теле животного направления: латеральное, т. е. вбок (вправо и влево), и медиальное, т. е. к средней сагиттальной плоскости.
Как показано на рисунке 1, проксимальная бедренная кость собак и киномолгус-макак более близка к грубой и микронатомии проксимальной бедра человека, чем к кроликам, крысам и мышам, и поэтому они, вероятно, являются лучшими моделями для изучения физиологии проксимального человека бедра и тазобедренного сустава. Несмотря на то, что механические нагрузки в проксимальном бедре различаются между людьми и четвероногими, а уменьшенная физическая активность лабораторных животных влияет на биомеханические исследования, анатомические и структурные сходства проксимального бедра и тазобедренного сустава между людьми, собаками и нечеловеческими приматами позволяют проводить биомедицинские исследования в лабораторных условиях с использованием собак и нечеловеческих приматов.
Метамерия - metameria (meta - за, после, meros - часть), или сегментация (сегмент - отрезок), заключается в том, что тело может быть разделено сегментными плоскостями на определенное (но не произвольное) число сравнительно одинаковых по строению частей - метамеров, или сегментов. Метамеры следуют один за другим вдоль оси тела: у животных - спереди назад, а у человека - сверху вниз. На скелете такими мета-мерами являются позвонки с соответствующими парами ребер.
Однако следует помнить, что масса кости, грубая и микроанатомия оптимально спроектированы таким образом, чтобы обеспечить максимальную безопасность скелета при естественных действиях, которые уникальны для каждого вида. Наше исследование изменений скелета среди лабораторных животных показывает, что абсолютные значения, зарегистрированные для параметров костей, не могут быть непосредственно сопоставлены между видами. Наши данные не позволяют сделать вывод о том, что на основе, например, более высокой минеральной плотности костной ткани или более хорошо связанной губчатой структуры, скелет одного вида превосходит скелет у других.
Б. Существует еще одна закономерность строения тела: вдоль спины в позвоночном канале располагается спинной мозг (мозговая трубка), а вентрально от позвоночного столба размещается кишечная трубка со всеми ее производными (внутренностями). В результате такого построения, обусловленного действием силы тяжести, на теле намечаются еще два направления: дорсальное, т. е. в сторону спины (dorsum - спина), и ему противоположное - вентральное, т. е. в сторону живота (venter - брюхо). У животных эти направления обозначаются также терминами вверх и вниз, а у человека - назад и вперед.
Грубая и микроанатомическая структура поясничных позвонков значительно варьируется среди обычных лабораторных видов. Поскольку минеральная плотность костей была довольно схожей среди всех видов, мы считаем, что для измерения прочности кости на поясничном отделе позвоночника следует использовать измерения минеральной плотности объемной кости и структуры губчатой кости. Поскольку механические нагрузки на горизонтальный позвоночник в четвероногих отличаются от механических нагрузок у людей, при которых позвоночник сохраняет вертикальное положение, биомеханические результаты и данные, описывающие структурные изменения костей в позвоночнике лабораторных животных, требуют тщательной рационализации при экстраполяции на биомеханику в людях.
Скелет конечностей представлен двумя парами конечностей - грудными, или передними (у человека - верхними), и тазовыми, или задними (у человека - нижними). В скелете конечностей четко выражена только одна из рассмотренных закономерностей - антимерия. Поскольку конечности - парные органы (правые и левые), все элементы их построены асимметрично.
Скелет хордовых животных
Тем не менее, модели крыс могут быть успешно использованы для предоставления ценной информации о физиологии длинных костей и позвоночника, а различные модели крыс и лабораторные условия могут быть смоделированы в дальнейшем, чтобы лучше напоминать последствия остеопороза человека на скелете.
Несмотря на то, что механические силы обеспечивают критические сигналы для моделирования костей и ремоделирования на протяжении всего скелета, биомеханика челюстей уникальна и отличается от биогенной кости в осевом скелете. Во время кусания в челюстях возникает сложная картина стресса и деформаций. Диапазон и распределение механических нагрузок варьируется среди видов, но всегда зависит от характера применяемых нагрузок и свойств материала и геометрии челюстей. Окклюзионная сила во время кусания переносится с зубов на губчатую и кортикальную кость челюстей; хотя объем трабекулярной кости значительно ниже, чем у кортикальной кости, губчатая кость, окружающая гнездо зуба, играет ключевую роль в распределении зубов и распределении нагрузки.
Метамерия в конечностях позвоночных наблюдалась в далеком прошлом; она еще заметна у низших животных и сильно затушевана или совершенно отсутствует у наземных (следы метамерии у них сохранились в дистальных отделах конечностей - лапах).


Более того, трабекулярное структурное изменение отражает хрупкость костной ткани более непосредственно, чем уменьшение объема кости; поэтому крайне важно использовать модели животных, чьи челюсти отражают распределение губчатой кости, подобное таковому человеческой челюсти.
Возрастные изменения скелета
Многочисленные исследователи подчеркивали отсутствие моделей на животных как серьезное препятствие для поддержки стоматологических исследований, связанных с потерей костной ткани в челюстях, которая известна своим сложным патогенезом и этиологией. Атрибуты идеальной модели для стоматологических исследований будут включать анатомические и физиологические особенности, сравнимые с таковыми у людей, прогрессирование системной скелетной и черепно-лицевой болезни, сходные с состояниями человека, и возможность изучения как системных, так и локальных факторов.


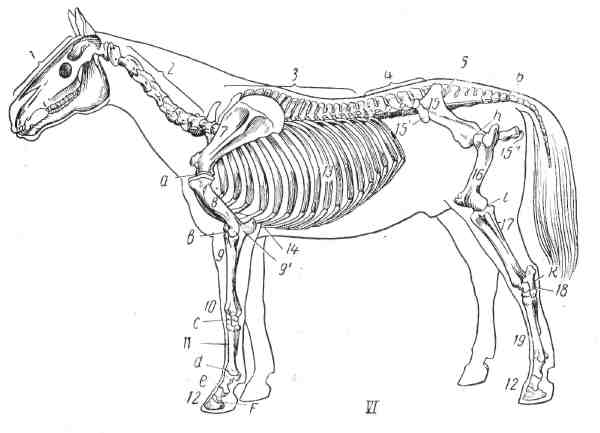
Рис. 5. Скелет: I- собаки; II- свиньи; III - коровы; IV - овцы; V - козы; VI - лошади.
1 - череп; 2-шейные позвонки; 3 - грудные позвонки; 4 - поясничные позвонки; 5 - крестцовая кость; 6 - хвостовые позвонки; 7 - лопатка; 8 - плечевая кость; 9 - кости предплечья (лучевая кость и 9′-локтевая кость); 10-кости запястья; 11-кости пясти; 12-кости пальцев-фаланги; 13-ребра; 14-грудная кость; 15-тазовая кость; 15′ -маклок подвздошной кости; 15″ - седалищный бугор; 16 - бедренная кость; 17 - большая берцовая кость; 18- кости заплюсны; 19 - кости плюсны. Суставы: a – плечевой, b – локтевой, c – запястный, d- путовый, e - венечный, f - копытный, h- тазобедренный, i - коленный, k - заплюсневый (на III, IV и V I).
Существующие модели животных имеют свои преимущества и недостатки. Грызуны и кролики легко доступны и могут быть использованы для ответа на некоторые основные вопросы. Однако они имеют существенные физиологические и анатомические отличия от людей и не представляют хорошей модели для стоматологических исследований, главным образом из-за присутствия резцов и относительно небольших областей с губчатой костью. Из всех видов, которые мы исследовали, кролики, по-видимому, являются наименее желательной моделью для изучения пероральной механики и патофизиологии для экстраполяции данных людям, потому что резцы и коренные зубы кроликов глубоко внедрены в костные части нижней челюсти, которая содержит очень маленькие участки губчатой кости.
На парных конечностях (грудных и тазовых) различают скелет поясов (грудного и тазового) и скелет свободных конечностей. В скелете поясов конечностей первоначально было по три пары костей.
Грудной, или плечевой, пояс в полном виде состоит из лопатки, ключицы и коракоидной кости; так устроен плечевой пояс у птиц, в то время как у домашних млекопитающих животных из перечисленных трех костей имеется лишь одна лопатка. Верхний конец лопатки направлен дорсо-каудально, а нижний конец - вентро-краниально, причем он участвует в образовании плечевого сустава. Рудимент ключицы встречается у собаки и кошки, а от коракоидной кости у всех млекопитающих животных сохранился лишь небольшой отросток на бугре лопатки с медиальной стороны! (У человека, помимо лопатки, имеется ключица, которая лежит в сегментной плоскости и обеспечивает возможность хватательных движений.)
Кроме того, присутствие резцов и небольшой зоны губчатой кости значительно ограничивает использование грызунов для изучения физиологии сухожилий. Исходя из результатов текущего исследования, киномолгусные макаки и собаки, по-видимому, являются образцами выбора для доклинических исследований болезни пародонта и зубных эффектов, связанных с потерей костной ткани, поскольку костная структура челюстей и анатомия моляров имитируют человеческую анатомию и физиология; однако другие виды, включая свиней, могут также использоваться в стоматологических исследованиях.
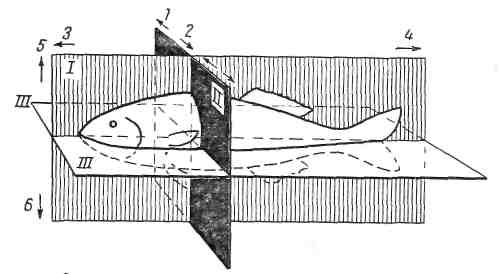
Рис. 6. А. Плоскости и направления в теле: I - разрез в средней сагиттальной плоскости; II - разрез в сегментальной плоскости; III - разрез во фронтальной плоскости. Направления: 1 - латеральное; 2 - медиальное; 3 - краниальное; 4 - каудальное; 5 - дорсальное; 6 - вентральное.
Размер и форма нижнечелюстного тела функционально связаны с биомеханикой жевания и кусания и, следовательно, специфичны для видов. Помимо костной массы и структуры, между лабораторными животными и людьми существуют другие соответствующие различия в форме и функции, включая силы, действующие на нижнюю челюсть во время жевания, и эти различия следует взвешивать при экстраполяции данных исследований животных на людей.
В совокупности результаты этого исследования подчеркивают ключевые параметры грубой анатомии и микроанатомии проксимальной бедра, поясничного отдела позвоночника и нижней челюсти широко используемых лабораторных видов. Описанные скелетные характеристики и человеческая значимость свидетельствуют о том, что кролик, по-видимому, является наименее желательным видом для проведения доклинических исследований физиологии костной ткани из-за морфологии проксимальной бедра, включая очень короткую шейку бедренной кости и небольшое количество губчатой кости на шейке бедренной кости, тело позвонка, и нижней челюсти, тогда как циномологические макаки, вероятно, являются наиболее подходящими среди изучаемых нами видов.
В тазовом пояс е хорошо развиты все три тазовые кости: подвздошная, лонная и седалищная. Подвздошная кость направлена дорсо-краниально, она соединяется с крылом крестцовой кости, образуя крестцово-подвздошный сустав. Лонная кость лежит в сегментной плоскости, соединяя правую подвздошную кость с левой. Седалищная кость находится каудально от подвздошной. Все три кости участвуют в образовании тазобедренного сустава.
Однако модели грызунов, особенно модели крыс, чрезвычайно полезны для проведения фундаментальных исследований скелета и представляют собой хорошую и доступную альтернативу собакам и нечеловеческим приматам. Несмотря на то, что прямые сопоставления между видами сложны и экстраполирование данных, полученных для человека, может быть сложным, мы считаем, что лучшее изображение анатомических и структурных характеристик скелетных участков в лабораторных видах, представленных здесь, поможет ученым в выборе подходящих моделей животных и скелета сайтов, которые лучше всего поддерживают их конкретные исследования.
В скелете свободных конечностей различают, считая сверху вниз: первые, вторые и третьи звенья. Первые звенья, непосредственно сочленяющиеся с поясами, состоят из одного луча, отсюда и его название стилоподий - stylopodium (stylos - столбик, греч. pous, podos - нога); на грудной конечности - это плечевая кость, а на тазовой - бедренная кость. Вторые звенья представлены двумя лучами на каждой конечности и поэтому именуются зейгоподием - zeugopo-dium (zeugos - пара, две); на грудной конечности - это лучевая и локтевая кости предплечья, а на тазовой - большая берцовая и малая берцовая кости голени.
На костях свободных отделов конечностей различают поверхности: латеральную и медиальную (как на туловище), переднюю - дорсальную, или краниальную, и заднюю - каудальную поверхность или: на грудной конечности - волярную - volaris, а на тазовой - плантарную - plantaris.
Третьи звенья образуют скелет передней и задней лапы - автоподий - autopodium (autos - он сам). Лапа грудной конечности соответствует кисти человека, а лапа тазовой конечности - стопе. Количество лучей в лапе у разных животных неодинаково. В каждой лапе, в свою очередь, различают: базиподий - basipodium - основной проксимальный (ближайший к туловищу) участок, метаподий - metapodium - средний участок иакроподий - acropodium (acros -самый крайний)- дистальный, или концевой, участок лапы.
Базиподий передней лапы, или запястье, представлен серией коротких костей, расположенных в два ряда, а в базиподий задней лапы, или в заплюсне, кости расположены в три ряда. Метаподий представлен на грудной конечности костями пясти, а на тазовой конечности - костями плюсны. Акроподий представлен на обеих конечностях костями пальцев, или фалангами,в количестве трех на каждом пальце: проксимальной (верхней), средней и дистальной (концевой). Дистальная, или концевая, фаланга называется копытной костью у лошади, копытцевой у жвачных и свиньи и ногтевой костьюу собаки.
Автоподий (лапа) служит для непосредственной опоры о землю или хватательным органом. В зависимости от особенностей функции (опоры или хватания) лапа устроена неодинаково в ряду животных, ее функция оказывает влияние на строение зейгоподия, в котором обе кости или хорошо развиты, или они срастаются друг с другом, или развита хорошо только одна кость.
Звенья свободных отделов конечностей имеют неодинаковое направление на грудной и тазовой конечностях.

Рис. 6 Б. Схема распилов тела животного через общий и частные центры тяжести. Показано на осевом скелете (по В. Я. Бровару, 1960): (сравн. с рис. 5):
1- распил через общий центр тела животного, на переднюю и заднюю половины; 2 - распилы через центр краниальной половины тела на первую и вторую четверти тела; 3 - распил через центр каудальной половины тела на третью и четвертую четверти тела; 4, 5, 6 и 7 - распилы каждой четверти на восьмые части тела. В результате распилов через центры обособляются: голова, шея, крестец и хвост. В плоскости общего центра тела (1) лежит печень; в плоскости центра второй четверти (5) лежит сердце; в плоскости центра третьей четверти (6) лежат почки.
Плечевая кость своим проксимальным (верхним) концом направлена дорсо – краниально (вверх и вперед,) где совместно с лопаткой образует плечевой сустав, а дистальным (или нижним) концом обращена каудо-вентрально (назад и вниз) и совместно с костями предплечья образует локтевой сустав.
Бедренная кость проксимальным концом обращена, наоборот, дорсо-каудально (вверх и сзади), где совместно с тазовой костью образует тазобедренный сустав, дистальным же концом направлена кранио-вентрально (вперед и вниз) и с костями голени образует коленный сустав, в котором спереди еще имеется коленная чашка. Кости предплечья ниже локтевого сустава стоят почти вертикально; дистально они образуют запястный сустав. Кости голени, напротив, лежат косо: проксимальный, или коленный, их конец направлен дорсо-краниально (вперед и вверх), а дистальный или заплюсневый, - вентро-каудально (вниз и назад). Он образует заплюсневый, или скакательный, сустав. Кости пясти и соответственно плюсны стоят более или менее вертикально, образуя пястно (соответственно плюснево-)-фаланговый сустав. Угол этого сустава прп стоянии копытного животного открыт дорсально, т. е. вперед, на обеих конечностях, так как дистальный (нижний) конец пальца направлен вперед.
В. Деление скелета, а следовательно, и тела животного, на вышеописанные части также закономерно и обусловлено действием сил тяжести (В. Я. Бровар). Это доказывается следующим образом. Труп животного, которому придано естественное «стоячее» положение, замораживают и затем распиливают на восемь частей, сначала в плоскости общего центра тяжести на переднюю и заднюю половины, а затем каждую половину распиливают также на переднюю и заднюю части (четверти) в плоскостях их центров. Каждую четверть снова распиливают в плоскости их центров тяжестей на передние и задние (т. е. уже восьмые) части. В результате получается, что первый распил прошел через печень, второй распил в передней половине тела отделил шею от грудной клетки, а на конечности-прошел через плечевой сустав; в задней же половине тела отделилась поясница от крестца, а в конечности распил прошел через коленный сустав. В результате третьих распилов в первой четверти отделилась голова от шеи, во второй четверти разрез прошел через сердце, а на конечности - через локтевой и запястный суставы, в третьей четверти разрез прошел через почки с толстой кишкой и в четвертой четверти крестец отделился от хвоста, а на конечности разрез прошел через тазобедренный сустав и скакательный.
Кроме перечисленных выше костей скелета, в организме существуют кости, не относящиеся собственно к скелету. Такими костями являются: у свиньи - хоботная кость - os rostri; у собаки кость головки полового члена - os penis! у рогатого скота - сердечные кости - ossa cordis. В других случаях кости оказываются окостеневшими участками сухожилий, как, например, сесамовидные кости проксимальной и дистальной фаланги - ossa sesamoidea; коленная чашка - patella - или тонкие длинные кости окостеневших сухожилий у птиц (например, у индеек).
АНАТОМИЯ ДОМАШНИХ ЖИВОТНЫХ
ПЛОСКОСТИ ТЕЛА И ТЕРМИНЫ ДЛЯ ОБОЗНАЧЕНИЯ РАСПОЛОЖЕНИЯ ОРГАНА
Для определения расположения органов и частей тело животного расчленяют тремя воображаемыми взаимоперпендикулярными плоскостями – сагиттальной, сегментальной и фронтальной (рис.1).
Срединная сагиттальная (медианная) плоскость проводится вертикально вдоль середины тела животного от рта до кончика хвоста и рассекающая его на две симметричные половины. Направление в теле животного к срединной плоскости называется медиальным, а от нее – латеральным (lateralis - боковой).
Рис.1. Плоскости и направления в теле животного
Плоскости:
I – сегментальная;
II - сагиттальная;
III – фронтальная.
Направления:
1 – краниальное;
2 – каудальное;
3 – дорсальное;
4 – вентральное;
5 – медиальное;
6 – латеральное;
7 – ростральное (оральное);
8 – аборальное;
9 – проксимальное;
10 – дистальвое;
11 – дорсальное
(спинковое, тыльное);
12 – пальмарное;
13 - плантарное.
Сегментальную плоскость проводят вертикально поперек тела животного. Направление от нее в сторону головы называется краниальным (cranium - череп), в сторону хвоста – каудальным (cauda - хвост). На голове, где все краниальное, различают направление к носу – назальное или хоботку – ростральное и противоположное ему-каудальное.
Фронтальную плоскость (frons - лоб) проводят горизонтально вдоль тела животного (при горизонтально вытянутой голове), т. е. параллельно лбу. Направление в данной плоскости в сторону спины называется дорсальным (dorsum - спина), к животу – вентральным (venter - живот).
Для определения положения участков конечностей существуют термины проксимальный (proximus - ближайший) – более близкое положение к осевой части тела и дистальный {distalus - удаленный) – более отдаленное положение от осевой части тела. Для обозначения передней поверхности конечностей приняты термины краниальный или дорсальный (для лапы), а для задней поверхности – каудальный, а также пальмарный или волярный (palma, vola - ладонь) – для кисти и плантарный (planta - стопа) – для стопы.
ОТДЕЛЫ И ОБЛАСТИ ТЕЛА ЖИВОТНОГО И ИХ КОСТНАЯ ОСНОВА
 |
Тело животных делят на осевую часть и конечности. Начиная с амфибий, у животных осевую часть тела делят на голову, шею, туловище и хвост. Шея, туловище и хвост составляют ствол тела. Каждая из частей тела разделена на отделы и области (рис.2). Основой их в большинстве случаев являются кости скелета, имеющие те же названия, что и области.
Рис. 2 Области тела крупного рогатого скота
1 - лобная; 2 - затылочная; 3 - теменная; 4 - височная; 5 - околоушная; 6 - ушной раковины; 7 - носовая; 8 - области верхней и нижней губ; 9 - подбородочная; 10 - щечная; 11 - межчелюстная; 12 - подглазничная; 13 - скуловая; 14 - область глаза; 15 - большой жевательной мышцы; 16 - верхняя шейная; 17 – боковая шейная; 18 - нижняя шейная; 19 - холки; 20 - спины; 21 - реберная; 22 - предгрудинная; 23 - грудинная: 24 - поясничная: 25 - подреберья; 26 - мечевидного хряща; 27 - околопоясничyая (голодная) ямка; 28 - боковая область; 29 - паховая; 30 - пупочная; 31 - лонная; 32 - маклок; 33 – крестцовая; 34 - ягодичная; 35 - корень хвоста; 36 - седалищная область; 37 - лопатка; 38 - плечо; 39 - предплечье; 40 - кисть; 41 - запястье; 42 - пясть; 43 - пальцы; 44 - бедро; 45 - голень; 46 - стопа; 47 - заплюсна; 48 - плюсна.
Голова (лат. caput, греч. cephale) делится на череп (мозговой отдел) и лицо (лицевой отдел). Череп (cranium) представлен областями: затылочной (затылок), теменной (темя), лобной (лоб) с областью рога у рогатого скота, височной (висок) и околоушной (ухо) с областью ушной раковины. На лице (facies) различают области: глазничную (глаза) с областями верхнего и нижнего век, подглазничную, скуловую с областью большой жевательной мышцы (у лошади – ганаши), межчелюстную, подбородочную, носовую (нос) с областью ноздрей, ротовую (рот), в состав которой входят области верхней и нижней губ и щеки. Над верхней губой (в области ноздрей) находится носовое зеркальце, у крупных жвачных оно распространяется на область верхней губы и становится носогубным.
Шея
Шея (cervix, collum) простирается от затылочной области до лопатки и делится на области: верхнюю шейную, лежащую над телами шейных позвонков; боковую шейную (область плечеголовной мышцы), идущую вдоль тел позвонков; нижнюю шейную, вдоль которой тянется яремный желоб, а также гортанную и трахейную (на вентральной ее стороне). У копытных шея сравнительно длинная в связи с необходимостью питания подножным кормом. Самая длинная шея у быстроаллюрных лошадей. Самая короткая – у свиньи.
Туловище
Туловище (truncus) состоит из грудного, брюшного и тазового отделов.
Грудной отдел включает области холки, спины, боковые реберные, предгрудинную и грудинную. Он прочный и подвижный. В каудальном направлении прочность уменьшается, а подвижность нарастает благодаря особенностям их соединения. Костной основой холки и спины являются грудные позвонки. В области холки у них наиболее высокие остистые отростки. Чем выше и длиннее холка, тем больше площадь прикрепления мышц позвоночника и пояса грудной конечности, тем размашистее и более упруги движения. Между длиной холки и спины существует обратная зависимость. Самая длинная холка и короткая спина у лошади, у свиньи – наоборот.
Брюшной отдел включает поясницу (lumbus), живот (abdomen), или брюхо (venter), поэтому еще называется пояснично-брюшным отделом. Поясница – продолжение спины до крестцовой области. Ее основой являются поясничные позвонки. Живот имеет мягкие стенки и делится на ряд областей: правого и левого подреберий, мечевидного хряща; парную боковую (подвздошную) с голодной ямкой, примыкающую снизу к пояснице, спереди – к последнему ребру, а сзади – переходит в паховую область; пупочную, лежащую снизу живота позади области мечевидного хряща и впереди от лонной области. На вентральной поверхности областей мечевидного хряща, пупочной и лонной у самок расположены молочные железы. У лошади наиболее короткая поясница и менее обширный брюшной отдел. У свиньи и рогатого скота поясница более длинная. Наиболее объемистый брюшной отдел у жвачных.
Тазовый отдел (pelvis) делится на области: крестцовую, ягодичную, включая маклок, седалищную и промежностную с примыкающей мошоночной областью. В хвосте (cauda) различают корень, тело и кончик. Области крестцовая, две ягодичных и корень хвоста у лошади образуют круп.
Конечности (membra) подразделяются на грудные (передние) и тазовые (задние). Состоят они из поясов, которыми соединяются со стволовой частью тела, и свободных конечностей. Свободные конечности делятся на основной поддерживающий столб и лапу. Грудная конечность состоит из плечевого пояса, плеча, предплечья и кисти.
Области плечевого пояса и плеча примыкают к боковой грудной области. Костной основой плечевого пояса у копытных является лопатка, поэтому его часто называют областью лопатки. Плечо (brachium) расположено ниже плечевого пояса, имеет форму треугольника. Костной основой является плечевая кость. Предплечье (antebrachium) находится вне кожного туловищного мешка. Его костная основа – лучевая и локтевая кости. Кисть (manus) состоит из запястья (carpus), пясти (metacarpus) и пальцев (digiti). У животных разных видов их бывает от 1 до 5. Каждый палец (кроме первого) состоит из трех фаланг: проксималыной, средней и дистальной (которые у копытных называются соответственно путовой, у лошади – бабка), венечной и копытцевой (у лошади – копытной).
Тазовая конечность состоит из тазового пояса, бедра, голени и стопы.
Область тазового пояса (таз) входит в состав осевой части тела в качестве ягодичной области. Костная основа – тазовая или безымянная кости. Область бедра (femur) расположена под тазом. Костная основа – бедренная кость. Область голени (crus) находится вне кожного туловищного мешка. Костная основа - большая и малая берцовые кости. Стопа (pes) состоит из заплюсны (tarsus), плюсны (metatarsus) и пальцев (digiti). Количество их, строение и названия у копытных такие же, как на кисти.
СОМАТИЧЕСКИЕ СИСТЕМЫ
Кожный покров, скелетная мускулатура и скелет, формируя собственно тело – сому животного,-объединяются в группу соматических систем организма.
Аппарат движения образован двумя системами: костной и мышечной. Кости, объединенные в скелет, представляют собой пассивную часть аппарата движения, являясь рычагами, на которые действуют прикрепленные к ним мышцы. Мышцы действуют только на кости, подвижно соединенные с помощью связок. Мышечная система – активная часть аппарата движения. Он обеспечивает движение организма, его перемещение в пространстве, поиск, захват и пережевывание пищи, нападение и защиту, дыхание, движения глаз, ушей и др. На его долю приходится от 40 до 60% массы организма. Он определяет форму тела животного (экстерьер), пропорции, обусловливая типовые особенности конституции, что имеет огромное практическое значение в зоотехнии, т. к. с особенностями экстерьера, типом конституции связаны выносливость, приспособляемость, способность к откорму, скороспелость, половая активность, жизнестойкость, и другие качества животных.
СКЕЛЕТ, СОЕДИНЕНИЕ КОСТЕЙ СКЕЛЕТА (ОСТЕОЛОГИЯ)
Общая характеристика и значение скелета.
Скелет (греч. skeleton - иссохший, мумия) образован костями и хрящами, соединенными между собой соединительной, хрящевой или костной тканями. Скелет млекопитающих называется внутренним, т. к. он расположен под кожей и покрыт слоем мышц. Он является твердой основой тела и служит футляром для головного, спинного и костного мозга, для сердца, легких и других органов. Упругость и рессорные свойства скелета обеспечивают плавность движений, предохраняют мягкие органы от толчков и сотрясений. Скелет участвует в минеральном обмене. В нем содержатся большие запасы солей кальция, фосфора и др. веществ. Скелет – наиболее точный показатель степени развития и возраста животного. Многие прощупываемые кости являются постоянными ориентирами при проведении зоотехнических измерений животного.
ДЕЛЕНИЕ СКЕЛЕТА
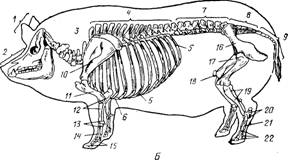
Скелет делится на осевой и скелет конечностей (периферический) (рис.3).
Осевой скелет включает скелет головы, шеи, туловища и хвоста. Скелет туловища состоит из скелета грудной клетки, поясницы и крестца. Периферический скелет образован костями поясов и свободных конечностей. Количество костей у животных разных видов, пород и даже особей неодинаково. Масса скелета составляет у взрослого животного от 6% (свиньи) до 12-15% (лошадь, бык). У новорожденного телят – до 20%, а у поросят – до 30%. от массы тела. У новорожденных более развит периферический скелет. На его долю приходится 60-65% массы всего скелета, а на долю осевого 35-40%. После рождения активнее растет, особенно в молочный период, осевой скелет и у 8-10-месячного теленка отношения этих отделов скелета выравниваются, а затем осевой начинает преобладать: в 18 месяцев у крупного рогатого скота он составляет 53-55%. У свиньи масса осевого и периферического скелета примерно одинакова.
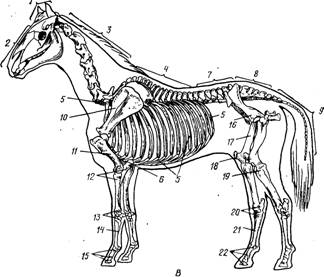 |
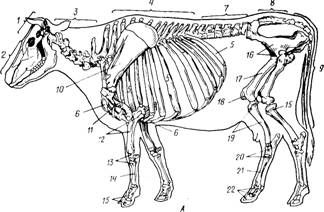 Рис.3 Скелет коровы (А), свиньи (Б),
Рис.3 Скелет коровы (А), свиньи (Б),
лошади (В)
Осевой скелет: 1- кости мозгового отдела (черепа): 3- кости лицевого отдела (лица); а- шейные позвонки; 4 - грудные позвонки; 5 - ребра; 6 - грудина; 7 - поясничные позвонки: 8 - крестцовая кость: 9 - хостовые позвонки (3,4,7,8,9 - позвоночник). Скелет конечностей; 10 - лопатка; 11 - плчевая кость; 12 - кости предплечья (лучевая и локтевая); 13 - кости запястья; 14 - кости пясти; 15 - кости пальцев (IS-15 – кости кисти); 16 - тазовая кость; П - бедренная кость: IS - коленная чашка; IS - кости голени (большая и малая берцовые); 30 - кости заплюсны: 31 - кости плюсны; 32 - кости пальцев (20-22 - кости стопы).
Форма и строение костей
Кость (лат. os) – орган костной системы. Как всякий орган, она имеет определенную форму и состоит из нескольких видов тканей. Форма костей определяется особенностями ее функционирования и положением в скелете. Различают длинные, короткие, плоские и смешанные кости.
Длинные кости бывают трубчатыми (многие кости конечностей) и дугообразными (ребра). Длина тех и других больше ширины и толщины. Длинные трубчатые кости напоминают по форме цилиндр с утолщенными концами. Средняя, более узкая часть кости называется телом – диафиз (греч. diaphysis), расширенные концы – эпифизы (epiphysis). Эти кости играют основную роль в статике и динамике, в кроветворной функции (содержат красный костный мозг).
Короткие кости обычно небольшой величины, их высота, ширина и толщина близки по размеру. Они часто выполняют рессорную функцию.
Плоские кости имеют большую поверхность (ширину и длину) при малой толщине (высоте). Обычно они служат стенками полостей, защищая помещенные в них органы (черепномозговая коробка) или это обширное поле для прикрепления мышц (лопатка).
Смешанные кости имеют сложную форму. Эти кости, как правило, непарные и размещаются по оси тела. (затылочная, клиновидная кости, позвонки). Парные смешанные кости несимметричны, например височная кость.
Строение кости
Основная ткань, образующая кость – пластинчатая костная. В состав кости входят также ретикулярная, рыхлая и плотная соединительные ткани, гиалиновый хрящ, кровь и эндотелий сосудов, нервные элементы.
Снаружи кость одета надкостницей, или периостом, за исключением места расположения суставного хряща. Наружный слой надкостницы фиброзный, образован соединительной тканью с большим количеством коллагеновых волокон; определяет ее прочность. Внутренний слой содержит недифференцированные клетки, которые могут преобразовываться в остеобласты и являются источником роста кости. Через надкостницу в кость проникают сосуды и нервы. Надкостница во многом определяет жизнеспособность кости. Кость, очищенная от надкостницы, погибает.
Под надкостницей залегает слой кости, образованный плотно уложенными костными пластинками. Это компактное вещество кости. В трубчатых костях в нем различают несколько зон. К надкостнице примыкает зона наружных генеральных пластинок толщиной 100-200 мкм. Она придает кости большую твердость. Затем следует наиболее широкая и важная в структурном отношении зона остеонов. Чем толще слой остеонов, тем лучше рессорные свойства кости. В этом слое между остеонами залегают вставочные пластинки – остатки старых разрушенных остеонов. У копытных в нем часто встречаются циркулярно-параллельные структуры, устойчивые к сопротивлению на изгиб. Не случайно они широко распространены в длинных трубчатых костях копытных, испытывающих большое давление. Толщина внутреннего слоя компактного вещества 200-300 мкм, образован он внутренними генеральными пластинками или же переходит в губчатое вещество кости.
Губчатое вещество представлено костными пластинками, которые не плотно примыкают друг к другу, а формируют сеть из костных перекладин (трабекул), в ячеях которой располагается красный костный мозг. Губчатое вещество особенно развито в эпифизах. Его перекладины располагаются не беспорядочно, а строго следуют линиям действующих сил (сжатия и растяжения).
В середине диафиза трубчатой кости имеется костная полость . Она образовалась в результате резорбции костной ткани остеокластами во время развития кости и заполнена желтым (жировым) костным мозгом.
Кость богата сосудами, которые образуют сеть в ее надкостнице, пронизывают всю толщину компактного вещества, находясь в центре каждого остеона, и разветвляются в костном мозге. В кости кроме сосудов остеонов имеются т. н. питательные сосуды (фолькмановы), прободащие кость перпендикулярно ее длине. Вокруг них не образуется концентрических костных пластинок. Особенно много таких сосудов около эпифизов. Нервы в кость проникают от надкостницы через те же отверстия, что и сосуды. Поверхность кости покрыта гиалиновым хрящом без надхрящницы. Толщина его 1-6 мм и прямо пропорциональна нагрузке на сустав.
Строение коротких, сложных и плоских костей такое же, как и трубчатых, с той лишь разницей, что у них обычно нет костных полостей. Исключение составляют некоторые плоские кости головы, в которых между пластинками компактного вещества имеются обширные пространства, заполненные воздухом – синусы или пазухи.
ФИЛОГЕНЕЗ СКЕЛЕТА
Развитие опорной системы в филогенезе животных шло двумя путями: образования наружного и внутреннего скелета. Наружный скелет закладывается в покровах тела (членистоногие). Внутренний скелет развивается под кожей и обычно прикрыт мышцами. О развитии внутреннего скелета можно говорить со времени появления хордовых. У примитивных хордовых (ланцетник) – хорда представляет собой опорную систему. С усложнением организации животных соединительнотканный скелет замещается хрящевым, а затем костным.
Филогенез стволового скелета
В филогенезе позвоночных раньше других элементов появляются позвонки. С усложнением организации, увеличением активности и разнообразия движений вокруг хорды развиваются не только дуги, но и тела позвонков. У хрящевых рыб скелет образован хрящом, иногда обызвествленным. Кроме верхних дуг под хордой у них развиваются нижние дуги. Концы верхних дуг каждого сегмента, сливаясь, образуют остистый отросток. Появляются тела позвонков. Хорда теряет значение опорного стержня. У костистых рыб происходит замещение хрящевого скелета костным. Появляются суставные отростки, которыми позвонки сочленяются друг с другом, чем обеспечивается прочность скелета при сохранении его подвижности. Осевой скелет разделяется на головной, туловищный с ребрами, охватывающими полость тела с органами, и сильно развитый хвостовой отдел – локомоторный.
Переход к наземному образу жизни приводит к развитию одних частей скелета и редукции других. Скелет туловища дифференцируется на шейный, грудной (спинной), поясничный и крестцовый отделы, частично редуцируется скелет хвоста, т. к. основная нагрузка при движении по земле падает на конечности. В грудном отделе в тесной связи с ребрами развивается грудина, формируется грудная клетка. У земноводных шейный и крестцовый отделы позвоночника имеют только по одному позвонку, поясничный отдел отсутствует. Ребра очень короткие, у многих срастаются с поперечными отростками позвонков. У рептилий шейный отдел удлиняется до восьми позвонков и приобретает большую подвижность. В грудном отделе 1-5 пар ребер соединены с грудиной – образуется грудная клетка. Поясничный отдел длинный, имеет ребра, величина которых уменьшается в каудальном направлении. Крестцовый отдел образован двумя позвонками, хвостовой отдел длинный, хорошо развит.
У млекопитающих, независимо от образа жизни, постоянное число шейных позвонков (7). Относительно постоянно число позвонков и в остальных отделах: 12-19 грудных, 5-7 поясничных, 3-9 крестцовых. Хвостовых позвонков насчитывают от 3 до 46. Позвонки, за исключением первых двух, соединены при помощи хрящевых дисков (менисков), связок и суставных отростков.
Поверхности тел шейных позвонков часто имеют выпукло-вогнутую форму – опистоцельные. В остальных отделах позвонки обычно плоские-платицельные. Ребра сохраняются только в грудном отделе. В пояснице они редуцируются и срастаются с поперечными отростками позвонков. В крестцовом отделе срастаются и позвонки, образуя крестцовую кость. Хвостовой отдел облегчен, его позвонки сильно редуцируются.
Филогенез скелета головы
Скелет головного конца тела развивается вокруг нервной трубки – осевой (мозговой) скелет головы и вокруг головной кишки – висцеральный. Осевой скелет головы представлен хрящевыми пластинками, окружающими нервную трубку снизу и с боков, крыша черепа перепончатая. Висцеральный скелет головы состоит из хрящевых жаберных дуг, связанных с аппаратом дыхания и пищеварения; челюстей нет. Развитие скелета головы шло путем объединения мозгового и висцерального скелетов и усложнения их строения в связи с развитием головного мозга, органов чувств (обоняния, зрения, слуха). Мозговой череп хрящевых рыб представляет собой сплошную хрящевую коробку, окружающую головной мозг. Висцеральный скелет образован хрящевыми жаберными дугами. Черепная коробка у костистых рыб сложного строения. Первичные кости формируют затылочную область, часть основания черепа, обонятельную и слуховую капсулы и стенку глазницы. Покровные кости покрывают первичную черепную коробку сверху, снизу и с боков. Висцеральный скелет представляет собой очень сложную систему рычагов, участвующих в хватательных, глотательных и дыхательных движениях. С черепной коробкой висцеральный скелет сочленяется при помощи подвеска (hyomandibulare), в результате чего образуется единый скелет головы.
С выходом на сушу, с резкой сменой среды обитания и образа жизни животных происходят значительные изменения в скелете головы: череп подвижно причленяется к шейному отделу; уменьшается количество костей черепа за счет их сращения; увеличивается его прочность. Смена типа дыхания (с жаберного на легочное) приводит к редукции жаберного аппарата и преобразованию его элементов в подъязычную и слуховые кости. Челюстной аппарат срастается с основанием черепа. В ряду наземных животных прослеживается постепенное усложнение. В черепе земноводных много хрящей, слуховая кость одна. Для черепа млекопитающих характерно уменьшение количества костей за счет их сращения (например, затылочная кость образуется путем сращения 4, а каменистая – 5 костей), в стирании граней между первичными и покровными (вторичными) костями, в мощном развитии обонятельной области и сложном звукопроводящем аппарате, в крупных размерах черепной коробки и др.
Филогенез скелета конечностей
Гипотеза о происхождении конечностей наземных животных на базе парных плавников рыб сейчас имеет широкое признание. Парные плавники в типе хордовых впервые появились у рыб. Костной основой парных плавников рыб служит система хрящевых и костных элементов. Тазовый пояс у рыб развит в меньшей степени. С выходом на сушу, на базе парных плавников развивается скелет конечностей, расчлененный на отделы, типичные для пятипалой конечности. Пояса конечностей состоят из 3 пар костей и укрепляются связью с осевым скелетом: плечевой пояс – с грудиной, тазовый с крестцом. Плечевой пояс состоит из коракоида, лопатки и ключицы, тазовый – из подвздошной, лонной и седалищной костей. Скелет свободных конечностей расчленен на 3 отдела: у передней конечности –это кости плеча, предплечья и кисти, у задней – бедра, голени и стопы.
Дальнейшие преобразования связаны с характером передвижения, его скоростью и маневренностью. У земноводных пояс грудных конечностей, причленяясь к грудине, не имеет жесткой связи с осевым скелетом. В поясе тазовых конечностей развита его вентральная часть. У рептилий в скелете поясов в равной степени развиты дорсальная и вентральная части.
Плечевой пояс млекопитающих редуцирован и состоит из двух или даже одной кости. У животных с развитыми отводящими движениями грудной конечности (например, кротов, летучих мышей, обезьян) развиты лопатка и ключица, а у животных с однообразными движениями (например, у копытных) – только лопатка. Тазовый пояс млекопитающих упрочен тем, что лонные и седалищные кости соединяются вентрально с соименными костями. Скелет свободных конечностей млекопитающих организован так, что тело животного поднято над землей. Приспособление же к различным видам передвижения (бег, лазанье, прыжки, полет, плавание) привело к сильной специализации конечностей у разных групп млекопитающих, что выражается в основном в изменении длины и угла наклона отдельных звеньев конечностей, формы суставных поверхностей, срастании костей и редукции пальцев.
Изменение в строении конечностей в филогенезе в связи с повышением специализации – приспособленностью к определенному виду движения подробнее всего изучено в ряду лошадей (). Предполагаемый предок лошади, совмещая в себе черты копытных и хищных, был размером с лисицу и имел пятипалые конечности с когтями, по форме приближающимися к копытцам. От разнообразных мягких движений по рыхлой земле с высокой растительностью (лес) до широких размашистых быстрых движений по сухим открытым пространствам (степь) происходило удлинение основного поддерживающего столба конечностей из-за раскрытия (увеличения) углов между ее звеньями. Лапа приподнималась, животное переходило от стопо - к пальцехождению. При этом наблюдалась постепенная редукция нефункционирующих пальцев. При переходе от пальце - к фаланго (копыто-) хождению в состав основного поддерживающего столба входит уже вся лапа, а редукция пальцев достигает максимума. У лошади полноразвитым сохраняется на конечности лишь III палец. У рогатого скота развитыми оказываются два пальца-III и IV.
Онтогенез скелета
В процессе индивидуального развития особи скелет проходит те же 3 стадии развития и в той же последовательности, что и в филогенезе: соединительнотканного, хрящевого и костного скелета.
Хорда как один из первых осевых органов закладывается в зародышевый период внутриутробного развития в результате дифференцировки энтодермы и мезодермы в период гаструляции. Вскоре вокруг нее образуется сегментированная мезодерма – сомиты, внутренняя часть которых – склеротомы, прилежащие к хорде, являются скелетогенными зачатками.
Соединительнотканная стадия. В области склеротомов идет активное размножение клеток, которые принимают вид мезенхимных, разрастаются вокруг хорды и превращаются в ее соединительнотканный футляр и в миосепты – соединительнотканные тяжи. Соединительнотканный скелет у млекопитающих существует очень короткое время, так как параллельно процессу обрастания хорды в перепончатом скелете происходит размножение мезенхимных клеток, особенно вокруг миосепт, и дифференцировка их в хрящевые.
Хрящевая стадия. Дифференцировка мезенхимных клеток в хрящевые начинается с шейной области. Первыми закладываются хрящевые дуги позвонков, которые образуются между хордой и спинным мозгом, обрастают спинной мозг сбоку и сверху, формируя его футляр. Смыкаясь между собой попарно над спинным мозгом, дуги формируют остистый отросток. В это же время из сгущений мезенхимных клеток, размножающихся в футляре хорды, развиваются хрящевые тела позвонков, а в миосептах – зачатки ребер и грудины. Замещениея соединительной ткани хрящом начинается у свиньи и овцы на 5-й, у лошади и крупного рогатого скота – на 6-й неделе эмбрионального развития. Затем в такой же последовательности, в какой шло образование хрящевого скелета, идет его окостенение.
В хрящевой закладке (модели) кости сосудов нет. С развитием кровеносной системы зародыша идет образование сосудов вокруг и внутри надхрящницы, в результате чего ее клетки начинают дифференцироваться не в хондробласты, а в остеобласты, т. е. она становится надкостницей – периостом. Остеобласты продуцируют межклеточное вещество и откладывают его поверх хрящевого зачатка кости. Формируется костная манжетка. Костная манжетка построена из грубоволокнистой костной ткани. Процесс образования и роста манжетки вокруг хрящевого зачатка называется окостенением.
Костная манжетка затрудняет питание хряща и начинает разрушаться. Первые очаги обызвествления и деструкции хряща обнаруживают в центре (диафизе) хрящевого зачатка. В очаг разрушающегося хряща от надкостницы проникают сосуды вместе с недифференцированными клетками. Здесь они размножаются и превращаются в костные клетки – возникает первый очаг (центр) окостенения. У каждой кости обычно несколько очагов окостенения (в позвонках копытных их насчитывают 5-6, в ребрах – 1-3).
В очаге окостенения остеокласты разрушают обызвествленный хрящ, бразуя лакуны и туннели, шириной 50-800 мкм. Остеобласты вырабатывают межклеточное вещество, которое откладывается по стенкам лакун и туннелей. Мезенхима, проникающая вместе с капиллярами, дает начало следующей генерации остеобластов, которые, откладывая межклеточное вещество в сторону стенок туннелей, замуровывают предыдущие генерации остеобластов – развиваются костные пластинки. Поскольку лакуны и туннели образуют сеть, костная ткань, выстилающая их, повторяет их форму и в целом напоминает губку, состоящую из переплетающихся костных тяжей, перекладин или трабекул. Из них формируется губчатое вещество кости. Образование кости внутри хрящевого зачатка на месте разрушенного хряща называется эндохондральным (энхондральным) окостенением.
Часть недифференцированных клеток, проникающих вместе с капиллярами в туннели и лакуны, превращается в клетки костного мозга, который заполняет пространства между костными трабекулами губчатого вещества.
Процесс энхондрального окостенения, начавшись в области диафиза, распространяется к концам зачатка – эпифизам. Параллельно с этим утолщается и нарастает костная манжетка. В таких условиях хрящевая ткань может расти только в продольном направлении. При этом хондробласты, размножаясь, выстраиваются друг над другом в виде клеточных колонок (монетных столбиков).
Закладка хрящевый моделей и их окостенение происходят быстро в тех участках тела, где потребность в опоре появляется очень рано. По срокам закладки и темпам дифференцировки костного скелета млекопитающих можно разделить на несколько групп. Копытные относятся к группе, у которой закладка и формирование очагов окостенения почти заканчиваются к моменту рождения, 90% кости образовано костной тканью. После рождения продолжается лишь рост этих очагов. Новорожденные таких животных активны, сразу могут самостоятельно передвигаться, следовать за матерью и добывать себе корм.
Первичные очаги окостенения в предплодный период отмечают в скелете туловища. У рогатого скота вначале окостеневают ребра. Окостенение позвонков начинается с атланта и распространяется в каудальном направлении. Тела окостеневают прежде всего у средних грудных позвонков. Во второй половине эмбрионального развития активна образуются остеоны, выделяются слои наружных и внутренних генеральных пластинок. В постнатальном онтогенезе идет нарастание новых слоев костной ткани до завершения роста животного, а также перестройка уже существующих остеонов.
Зона клеточных колонок постоянно нарастает со стороны эпифизов благодаря дифференцировке хрящевых клеток от надхрящницы. Со стороны же диафиза отмечается постоянное разрушение хряща из-за нарушения его питания и изменения химизма ткани. До тех пор, пока эти процессы уравновешивают друг друга, кость растет в длину. Когда же скорость энхондрального окостенения становится больше, чем скорость роста метаэпифизарного хряща, он истончается и полностью исчезает. С этого времени прекращается линейный рост животного. В осевом скелете дольше всего сохраняются хрящи между эпифизами и телом позвонков, особенно в крестце.
В энхондральной кости рост кости в ширину начинается из диафиза и выражается в разрушении старых и образовании новых остеонов, в формировании костной полости. В перихондральной кости перестройка заключается в том, что грубоволокнистая костная ткань манжетки замещается пластинчатой костной тканью в виде остеонов, циркулярно-параллельных структур и генеральных пластинок, составляющих в совокупности компактное вещество кости. В процессе перестройки образуются вставочные пластинки. У крупного рогатого скота и свиньи осевой скелет начинает окостеневать в 3-4 года, а полностью процесс завершается в 5-7 лет, у лошади – в 4-5 лет, у овцы – в 3-4 года.
Развитие черепа
Начало осевому черепу дают 7-9 сомитов. Вокруг конечного участка хорды склеротомы этих сомитов образуют сплошную перепончатую пластинку без следов сегментации. Она распространяется вперед (прехордально) и охватывает снизу и с боков мозговые пузыри, слуховые и обонятельные капсулы и глазные бокалы. Замещение соединительнотканного осевого черепа хрящевым начинается около переднего конца хорды под основанием головного мозга. Здесь закладывается пара околохордовых (парохордалии) хрящей. Дальше в оральном направлении закладываются две хрящевые балки или трабекулы. Т. к. они лежат впереди хорды, этот отдел осевого черепа называется прехордальным. Трабекулы и парахордалии, разрастаясь, сливаются вместе, образую основную хрящевую пластинку. В оральной части вдоль основной хрящевой пластинки закладывается хрящевая носовая перегородка, по обе стороны от которой развиваются носовые раковины. Затем хрящи замещаются первичными, или примордиальными, костями. Первичными костями осевого черепа являются затылочная, клиновидная, каменистая и решетчатая, образующие дно, переднюю и заднюю стенки черепномозговой полости, а также носовая перегородка и раковины. Остальные кости вторичные, кожные, или покровные, т. к. возникают из мезенхимы, минуя хрящевую стадию. Это теменные, межтеменные, лобные, височные (чешуя), формирующие крышу и боковые стенки черепномозговой полости.
Параллельно развитию осевого черепа идет преобразование висцерального скелета головы. Большая часть зачатков висцеральных дуг подвергается полной редукции, а часть их материала идет на образование слуховых косточек, подъязычной кости и хрящей гортани. Основная же масса костей висцерального скелета – вторичные, покровные. Осевой и висцеральный скелет головы млекопитающих настолько тесно связывается друг с другом, что кости одного входят в состав другого. Поэтому череп млекопитающих делят на мозговой отдел (собственно череп), являющийся вместилищем мозга, и лицевой отдел (лицо), образующий стенки носовой и ротовой полостей. В плодный период определяется форма черепа, характерная для вида и породы. Роднички – неокостеневшие участки – закрываются плотной соединительной тканью или хрящом.
Развитие конечностей
Конечности у млекопитающих закладываются в виде выростов шейно-грудных и пояснично-крестцовых сомитов. У КРС это происходит на 3-й неделе. Сегментация их не выражена. Закладки имеют вид скоплений мезенхимы, которые быстро увеличиваются в длину, превращаясь в лопастевидные выросты. Сначала эти выросты подразделяются на два звена: закладку поясов и свободных конечностей, не расчлененных на отделы и кости. Затем из сгущений мезенхимы дифференцируются соединительнотканные и хрящевые закладки костей. В процессе дифференцировки скелет конечностей проходит те же три стадии, что и стволовой скелет, но с некоторым отставанием. Окостенение конечностей у плода теленка начинается на 8-9-й неделе и протекает аналогично со стволовым скелетом. Многие выросты костей – апофизы. имеют собственные очаги окостенения. В процессе окостенения в трубчатых костях образуется губчатое и компактное вещество. Перестройка из центра кости распространяется на ее периферию. При этом в области диафиза благодаря деятельности остеокластов почти полностью исчезает губчатое вещество, сохраняясь лишь в эпифизах. Увеличивается костная полость. Красный костный мозг в ней становится желтым.
Слои компактного вещества становятся заметны в течение первых месяцев жизни. Степень его развития зависит от вида животного. У копытных в нем хорошо развиты генеральные пластинки и циркулярно-параллельные структуры, у хищных преобладают остеоны. Это связано с различиями в функциональных нагрузках костей, особенно конечностей. У копытных они приспособлены к прямолинейному движению и удержанию массивного тела, у хищных – к более легкому телу и разнообразным движениям.
В конечностях очаги окостенения появляются в костях поясов, распространяясь затем в дистальном направлении. Окончательное окостенение (синостоз) прежде всего происходит в дистальных звеньях. Так, у крупного рогатого скота окостенение дистальных звеньев конечности (плюсны и пясти) завершается к 2-2,5 годам, к 3-3,5 годам окостеневают все кости свободной конечности, а кости тазового пояса – лишь к 7 годам.
Возрастные изменения скелета
В связи с разными сроками закладки, скорости роста и окостенения костей скелета на протяжении онтогенеза происходит изменение пропорций тела. В период эмбрионального развития кости растут с разной интенсивностью. У копытных в первую половину интенсивнее растет осевой скелет, во вторую – скелет конечностей. Так, у 2-месячных плодов телят осевой скелет составляет 77%, скелет конечностей – 23%, а к рождению – 39 и 61%. По данным, у мериноса от времени хрящевой закладки (1-месячный эмбрион) до рождения скелет тазовой конечности с поясом увеличивается в 200 раз, грудной конечности – в 181 раз, таз – в 74 раза, позвоночник – в 30 раз, череп – в 24 раза. После рождения на смену усиленному росту периферического скелета приходит линейный рост осевого скелета.
В постнатальном онтогенезе скелет растет с меньшей скоростью, чем мышцы и многие внутренние органы, поэтому его относительная масса снижается в 2 раза. В процессе роста и дифференцировки костей увеличивается их прочность, что связано с увеличением число остеонов на единицу площади. От рождения до взрослого состояния толщина компактного вещества увеличивается в 3-4 раза, содержание в нем минеральных солей – в 5 раз, предельная нагрузка – в 3-4 раза, достигая у овец 280, у коров 1000 кг на 1 см2. Окончательной прочности кости крупного рогатого скота достигают уже к 12-месячному возрасту.
Xем крупнее животное, тем меньше у него запас прочности костей. У самцов кости толще, чем у самок, но недокорм сказывается на них сильнее. Улучшенные породы овец и свиней имеют более короткие и широкие кости конечностей. У скороспелых животных кости толще, чем у позднеспелых. Кости коров молочного типа лучше снабжаются кровью, а у коров мясного и мясо-молочного типов больше площадь компактного вещества кости и толщина стенки, что обусловливает большую прочность еее при нагрузках. Прочность кости на изгиб обусловливает структура остеонов. У свиней породы ландрас, например, прочность кости на изгиб выше, чем у крупной белой и сибирской северной пород, в связи с тем, что у ландрас более плотное расположение остеонов.
Из всех внешних условий на развитие скелета наибольшее влияние оказывает кормление и моцион. Улучшение кормления в период интенсивного роста костей ускоряет, недокорм угнетает их темпы роста, особенно в ширину, но не нарушает общих закономерностей роста скелета. У животных пастбищного содержания компактное вещество кости плотнее, в нем преобладают пластинчатые структуры, трабекулы губчатого вещества толще, равномернее по ширине и направлены строго по действию сил сжатия - растяжения. При стойловом и клеточном содержании животных замедляется рост и внутренняя перестройка костей, снижается их плотность и прочность по сравнению с выгульным, напольным содержанием и с животными, подвергающимися дозированному принудительному движению.
Добавление к рациону молодняка макро - и микроэлементов способствует образованию костей с более толстым компактным веществом и трабекулами и меньшей костной полостью. При недостатке минеральных веществ происходит деминерализация скелета, размягчение и резорбция позвонков, начиная с хвостовых.



